Faser av mitos och deras processer. Mitos. Stadier av mitos, deras varaktighet och egenskaper. Amitos. Mitos hos djur och växter
Interfas Är perioden mellan två celldelningar. I mellanfasen är kärnan kompakt, har ingen uttalad struktur, nukleolerna är tydligt synliga. Samlingen av interfaskromosomer är kromatin. Kromatin innehåller: DNA, proteiner och RNA i förhållandet 1: 1,3: 0,2, såväl som oorganiska joner. Kromatins struktur är förändrad och beror på cellens tillstånd.
Kromosomer i interfasen är inte synliga, så de studeras med elektronmikroskopiska och biokemiska metoder. Interfas innefattar tre steg: presyntes (G1), syntetisk (S) och postsyntetisk (G2). G-symbolen är en förkortning för engelska. gap - intervall; symbol S är en förkortning för engelska. syntes - syntes. Låt oss överväga dessa steg mer detaljerat.
Presyntetiskt stadium (G1). Kärnan i varje kromosom är en dubbelsträngad DNA-molekyl. Mängden DNA i cellen i det presyntetiska stadiet indikeras av symbolen 2c (från det engelska innehållet - innehåll). Cellen växer aktivt och fungerar normalt.
Syntetiskt stadium (S). Själv-duplicering, eller DNA-replikering, sker. I det här fallet dupliceras vissa delar av kromosomer tidigare, medan andra - senare, det vill säga DNA-replikering fortsätter asynkront. Parallellt finns en fördubbling av centrioler (om någon).
Postsyntetiskt stadium (G2). DNA-replikering slutar. Varje kromosom innehåller två dubbla DNA-molekyler, som är en exakt kopia av den ursprungliga DNA-molekylen. Mängden DNA i en cell vid postsyntesstadiet betecknas med symbolen 4c. Ämnen som är nödvändiga för celldelning syntetiseras. I slutet av interfasen slutar syntesprocesserna.
Mitosprocess
Prophase - den första fasen av mitos. Kromosomer spiralformas och blir synliga i ljusmikroskopet i form av tunna trådar. Centrioler (om några) avviker från cellens poler. I slutet av profas försvinner nukleolierna, kärnhöljet förstörs och kromosomerna tränger in i cytoplasman.
I profas ökar kärnans volym och kromosomer bildas som ett resultat av kromatinspiralisering. I slutet av profaset ser man att varje kromosom består av två kromatider. Nukleolierna och kärnhöljet löses gradvis upp och kromosomerna placeras slumpmässigt i cellens cytoplasma. Centriolerna avviker till cellens poler. En akromatinspindel med delning bildas, en del av filamenten som går från pol till pol, och en del av den fäster vid centromerer av kromosomer. Innehållet i genetiskt material i en cell förblir oförändrat (2n2xp).
Figur: 1. Schema för mitos i lökrotceller

Figur: 2. Schema för mitos i lökrotceller: 1-fas; 2,3 - profas; 4 - metafas; 5,6 - anafas; 7,8 - telofas; 9 - bildandet av två celler

Figur: 3. Mitos i cellerna i lökrotspetsen: a - interfas; b - profas; c - metafas; d - anafas; l, f - tidiga och sena telofaser
Metafas. Början av denna fas kallas prometafas. I prometafas är kromosomer ordnade ganska slumpmässigt i cytoplasman. En mitotisk apparat bildas, som inkluderar spindeln för delning och centrioler eller andra organ för organisation av mikrotubuli. I närvaro av centrioler kallas den mitotiska apparaten astral (i flercelliga djur) och i deras frånvaro anastral (i högre växter). Delningsspindeln (achromatinspindel) är ett system av tubulinmikrotubuli i en delande cell som säkerställer kromosomseparation. Delningsspindeln innehåller två typer av trådar: pol (stödjande) och kromosomal (dragning).
Efter bildandet av den mitotiska apparaten börjar kromosomer röra sig in i cellens ekvatorialplan; denna rörelse av kromosomer kallas metakinesis.
I metafas spiraliseras kromosomerna maximalt. Kromosomcentromererna ligger i cellens ekvatorialplan oberoende av varandra. Divisionsspindelns poltrådar sträcker sig från cellens poler till kromosomerna, och de kromosomala trådarna - från centromererna (kinetochores) - till polerna. Uppsättningen kromosomer i cellens ekvatorialplan bildar en metafasplatta.
Anafas. Separation av kromosomer i kromatider sker. Från detta ögonblick blir varje kromatid en oberoende kromatos med en kromatid, som är baserad på en DNA-molekyl. En-kromatidkromosomer i anafasgrupper avviker till cellens poler. När kromosomerna skiljer sig åt förkortas kromosomala mikrotubuli och polerna förlängs. I detta fall glider stolpen och kromosomtrådarna längs varandra.
Telofas. Klyvningsspindeln förstörs. Kromosomer vid cellens poler despiraliseras och kärnmembran bildas runt dem. Två kärnor bildas i cellen, genetiskt identiska med den ursprungliga kärnan. DNA-innehållet i dotterkärnor blir 2c.
Cytokines. Vid cytokinese uppdelas cytoplasman och membranet i dotterceller bildas. Hos djur inträffar cytokinese genom snörning av cellen. I växter uppträder cytokinese på olika sätt: i ekvatorialplanet bildas bubblor som smälter samman och bildar två parallella membran.
Vid detta slutar mitos och nästa mellanfas börjar.
Bland alla intressanta och ganska komplexa ämnen i biologin är det värt att lyfta fram två processer för celldelning i kroppen - meios och mitos... Först kan det verka som om dessa processer är desamma, eftersom celldelning i båda fallen inträffar, men i själva verket är det stor skillnad mellan dem. Först och främst måste du förstå mitos. Vad är denna process, vad är mitosfasen och vilken roll spelar de i människokroppen? Mer om detta och kommer att diskuteras i den här artikeln.
En komplex biologisk process, som åtföljs av celldelning och fördelningen av kromosomer mellan dessa celler - allt detta kan sägas om mitos. Tack vare honom fördelas kromosomer som innehåller DNA jämnt mellan kroppens dotterceller.
Det finns fyra huvudfaser i mitosprocessen. Alla är sammankopplade, eftersom faserna smidigt övergår från en till en annan. Förekomsten av mitos i naturen beror på det faktum att det är han som deltar i processen för uppdelning av alla celler, inklusive muskel, nerv och så vidare.
Kort om mellanfas
Innan mitten inträder går cellen, som är uppdelad, in i fasfasen, det vill säga den växer. Interfasens varaktighet kan ta mer än 90% av den totala tiden för cellaktivitet i normalt läge.
Interfasen är uppdelad i tre huvudperioder:
- fas Gl;
- S-fas;
- fas G2.
De äger rum alla i en specifik sekvens. Låt oss överväga var och en av dessa faser separat.
Interfas - huvudkomponenter (formel)
Fas G1
Denna period kännetecknas av förberedelsen av cellen för delning. Den ökar i volym för den ytterligare fasen av DNA-syntes.
S-fas
Detta är nästa steg i interfasprocessen, där celldelning sker i kroppen. Som regel sker syntesen av de flesta celler under en kort tidsperiod. Efter delning ökar inte cellerna i storlek, men den sista fasen börjar.
Fas G2
Det sista steget av interfas, under vilket celler fortsätter att syntetisera proteiner, samtidigt som de ökar i storlek. Under denna period innehåller cellen fortfarande nukleoli. I den sista delen av interfasen inträffar också duplicering av kromosomer, och ytan på kärnan vid denna tid är täckt med ett speciellt membran som har en skyddande funktion.
På en anteckning! I slutet av den tredje fasen inträffar mitos. Det innehåller också flera steg, varefter celldelning sker (denna process kallas cytokinese i medicin).
Stadier av mitos
Som nämnts tidigare är mitos uppdelad i fyra steg, men ibland kan det finnas fler. Nedan är de viktigaste.
Tabell. Beskrivning av mitos huvudfaser.
| Fasnamn, foto | Beskrivning |
|---|---|
| Under profas spiraliseras kromosomerna, vilket gör att de tar en vriden form (den är mer kompakt). Alla syntetiska processer i kroppens cell stoppas, så ribosomer produceras inte längre. |
| Många experter skiljer inte prometafas som en separat fas av mitos. Ofta kallas alla processer som äger rum i det som profas. Under denna period omsluter cytoplasman kromosomerna som rör sig fritt runt cellen till en viss punkt. |
| Nästa fas av mitos, som åtföljs av fördelningen av kondenserade kromosomer på ekvatorplanet. Under denna period förnyas mikrotubuli löpande. I metafas är kromosomerna placerade så att deras kinetokorer är i en annan riktning, det vill säga de riktas mot motsatta poler. |
| Denna fas av mitos åtföljs av separationen av kromatider från var och en av kromosomerna från varandra. Tillväxten av mikrorör slutar, de börjar nu demonteras. Anafas varar inte länge, men under denna tidsperiod lyckas cellerna spridas närmare olika poler i ungefär lika antal. |
| Detta är det sista steget under vilket kromosomdekondensation börjar. Eukaryota celler slutför sin uppdelning och ett speciellt skal bildas runt varje uppsättning mänskliga kromosomer. När den kontraktila ringen drar sig samman, är cytoplasman uppdelad (i medicin kallas denna process cytotomi). |





Viktig! Varaktigheten av den fullständiga mitosprocessen är som regel inte mer än 1,5-2 timmar. Varaktigheten kan variera beroende på vilken celltyp som delas. Externa faktorer som ljusförhållanden, temperatur och så vidare påverkar också processens varaktighet.
Vad är mitos biologiska roll?
Låt oss nu försöka förstå funktionerna i mitos och dess betydelse i den biologiska cykeln. För det första, det ger många viktiga processer i kroppen, inklusive embryonal utveckling.
Dessutom är mitos ansvarig för återställandet av vävnader och inre organ i kroppen efter olika typer av skador, vilket resulterar i regenerering. Under processen att dö av celler gradvis, men med hjälp av mitos bibehålls vävnadens strukturella integritet ständigt.
Mitos säkerställer bevarande av ett visst antal kromosomer (det motsvarar antalet kromosomer i moderns cell).
Video - Funktioner och typer av mitos
Föreläsning nummer 10
Antal timmar: 2
MITOS
1. Celllivscykel
2. Mitos. Stadier av mitos, deras varaktighet och egenskaper
3. Amitos. Endoreproduktion
1. Cellens livscykel
Cellerna i en multicellulär organism är extremt olika i sina funktioner. Celler har olika livslängd i enlighet med deras specialisering. Så nervceller efter avslutad embryogenes slutar dela och fungera under hela organismen. Celler av andra vävnader (benmärg, epidermis, tunntarmsepitel) i färd med att utföra sin funktion dör snabbt och ersätts av nya som ett resultat av celldelning.Celldelning är grunden för utveckling, tillväxt och reproduktion av organismer. Celldelning ger också självförnyelse av vävnader under hela kroppens livstid och återställer deras integritet efter skada. Det finns två sätt att dela upp somatiska celler: amitos och mitos... Indirekt celldelning (mitos) är främst utbredd. Reproduktion med mitos kallas asexuell reproduktion, vegetativ förökning eller kloning.
Celllivscykel (cellcykel) - detta är förekomsten av en cell från uppdelning till nästa uppdelning eller död. Varaktigheten av cellcykeln i multiplicerande celler är 10-50 timmar och beror på typen av celler, deras ålder, kroppens hormonbalans, temperatur och andra faktorer. Detaljerna i cellcykeln varierar mellan olika organismer. I encelliga organismer sammanfaller livscykeln med en individs liv. Vid kontinuerlig multiplicering av vävnadsceller sammanfaller cellcykeln med den mitotiska cykeln.
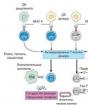
Mitotisk cykel -en uppsättning sekventiella och inbördes relaterade processer under beredningen av cellen för delning och delningsperioden (fig 1). I enlighet med ovanstående definition är den mitotiska cykeln uppdelad i mellanfasoch mitos (grekiska ”mitos” - tråd).
Interfas - perioden mellan två celldelningar - uppdelad i faser G 1,S och G 2 (Varaktigheten anges nedan, typiskt för växt- och djurceller.). När det gäller varaktighet utgör interfas det mesta av cellens mitotiska cykel. Mest varierande över tidG 1 och G 2 är perioder.
G 1 (från engelska.växa - växa, öka). Fasen varar 4–8 timmar. Denna fas börjar omedelbart efter cellbildning. I denna fas syntetiseras RNA och proteiner intensivt i cellen, aktiviteten hos enzymer som är involverade i DNA-syntes ökar. Om cellen inte delar sig vidare går den in i fasenG 0 - viloperiod. Med tanke på den vilande perioden kan cellcykeln pågå i veckor eller till och med månader (leverceller).
S (från engelska.syntes - syntes).Fasens varaktighet är 6–9 timmar Cellmassan fortsätter att öka och kromosomalt DNA dupliceras. Två spiraler av den gamla DNA-molekylen skiljer sig åt och blir var och en en matris för syntes av nya DNA-strängar. Som ett resultat innefattar var och en av de två dottermolekylerna nödvändigtvis en gammal helix och en ny. Icke desto mindre förblir kromosomerna enstaka i struktur, om än fördubblade i massa, eftersom de två kopiorna av varje kromosom (kromatider) fortfarande är anslutna till varandra längs hela sin längd. Efter fasens slut S av den mitotiska cykeln börjar cellen inte omedelbart dela sig.
G 2.I denna fas är processen för förberedelse för mitos slutförd i cellen: ATP ackumuleras, proteiner från akromatinspindeln syntetiseras, centrioler fördubblas. Cellmassan fortsätter att öka tills den är ungefär två gånger den initiala massan, och sedan börjar mitosen.
Figur: Mitotisk cykel: M - mitos, P - profas, Mf - metafas, OCH - anafas, T- telofas, G 1 - presyntesperiod, S - syntetisk period, G 2 - postsyntetisk
2. Mitos. Stadier av mitos, deras varaktighet och egenskaper. Mitos villkorligt uppdelad i fyra faser: profas, metafas, anafas och telofas.
Prophase. De två centriolerna börjar avvika mot kärnans motsatta poler. Kärnmembranet kollapsar; samtidigt kombineras speciella proteiner för att bilda mikrotubuli i form av trådar. Centriolerna, som nu är belägna vid motsatta poler i cellen, har en organiserande effekt på mikrotubuli, som följaktligen sträcker sig radiellt och bildar en struktur som ser ut som en asterblomma ("stjärna"). Andra trådar av mikrotubuli sträcker sig från en centriole till en annan och bildar en klyvningsspindel. Vid den här tiden spirar kromosomerna och tjocknar som ett resultat. De är tydligt synliga under ett ljusmikroskop, särskilt efter färgning. Att läsa genetisk information från DNA-molekyler blir omöjligt: \u200b\u200bRNA-syntes slutar, kärnan försvinner. I profas delas kromosomerna, men kromatiderna förblir fortfarande bundna parvis i centromerzonen. Centromerer har också en organiserande effekt på spindelns trådar, som nu sträcker sig från centriolen till centromeren och från den till den andra centriolen.
Metafas. I metafas når spiraliseringen av kromosomer ett maximum och de förkortade kromosomerna rusar till cellens ekvatorn, belägen på lika avstånd från polerna. Formad ekvator- eller metafasplatta. I detta stadium av mitos är kromosomernas struktur tydligt synlig, det är lätt att räkna dem och studera deras individuella egenskaper. Varje kromosom har en region av den primära förträngningen - centromeren, till vilken spindeltråden och axlarna är fästa vid mitos. I metafasstadiet består kromosomen av två kromatider som är anslutna till varandra endast i centromerregionen.

Figur: 1. Växtcell mitos. OCH - mellanfas;
B, C, D, D-
profas; E, F-metafas; 3, I - anafas; K, L, M-telofas
I anafas cytoplasmas viskositet minskar, centromererna separeras, och från detta ögonblick blir kromatiderna oberoende kromosomer. Spindelfilamenten fästa vid centromerer drar kromosomer till cellens poler, medan kromosomarmarna passivt följer centromeren. Således, i anafas, fördubblades kromatosomerna i kromosomerna i interfasen exakt till cellpolerna. För närvarande innehåller cellen två diploida uppsättningar kromosomer (4n4c).
Tabell 1. Mitotisk cykel och mitos
| Faser | Processen som äger rum i cellen |
|
| Interfas | Presyntetisk period (G1) | Proteinsyntes. RNA syntetiseras på despiraliserade DNA-molekyler |
| Syntetisk period (S) | DNA-syntes - självfördubbling av en DNA-molekyl. Konstruktionen av den andra kromatiden, i vilken den nybildade DNA-molekylen passerar: två-kromatidkromosomer erhålls |
|
| Postsyntetisk period (G2) | Proteinsyntes, energilagring, förberedelse för uppdelning |
|
| Faser mitos | Prophase | Bikromatidkromosomer spiraliserar, nukleoler upplöses, centrioler avviker, kärnhölje löses upp, klyvningspindeltrådar bildas |
| Metafas | Klyvningsspindelfilament fäster vid kromosomcentromerer, dikromatidkromosomer koncentrerar sig på cellekvatorn |
|
| Anafas | Centromerer delar, enkelkromatidkromosomer sträcks av spindelfilamenten till cellpolerna |
|
| Telofas | En-kromatidkromosomer despiraliseras, en kärna bildas, kärnhöljet återställs, en partition mellan celler börjar bildas vid ekvatorn, klyvningsspindelns trådar löses upp |
|
I telofas kromosomer varva ner, despiralisera. Ett kärnhölje bildas från cytoplasmas membranstrukturer. Vid denna tidpunkt återställs kärnan. Detta fullbordar uppdelningen av kärnan (karyokinese), då uppdelning av cellkroppen (eller cytokinese) inträffar. När djurceller delar sig uppträder en får på ytan i ekvatornas plan, vilket gradvis fördjupas och delas in i två halvor - dotterceller, som alla har en kärna. I växter sker uppdelning genom bildandet av den så kallade cellplattan som skiljer cytoplasman: den uppstår i spindelns ekvatorialområde och växer sedan i alla riktningar och når cellväggen (dvs. växer inifrån och ut). Cellplattan är bildad av material som levereras av det endoplasmatiska retikulumet. Sedan bildar var och en av dottercellerna ett cellmembran på sin sida och slutligen bildas cellulosacellväggar på båda sidor av plattan. Funktionerna i mitosförloppet hos djur och växter visas i tabell 2.
Tabell 2. Funktioner av mitos hos växter och djur
| Växtcell | Djurbur |
| Inga centrioles Stjärnor bildas inte En cellplatta bildas Ingen foder bildas under cytokenes Mitoser övervägande förekommer i meristems | Centrioles finns tillgängliga Stjärnor bildas Cellplattan är inte formad Med cytokinese bildas en får Mitoser uppstår i olika vävnader i kroppen |
Så från en cell bildas två dotterceller, i vilka ärftlig information exakt kopierar informationen i modercellen. Från och med den första mitotiska uppdelningen av ett befruktat ägg (zygote) innehåller alla dotterceller som bildats som ett resultat av mitos samma uppsättning kromosomer och samma gener. Följaktligen är mitos ett sätt för celldelning, som består i en exakt fördelning av genetiskt material mellan dotterceller. Som ett resultat av mitos får båda dottercellerna en diploid uppsättning kromosomer.
Hela mitosprocessen tar i de flesta fall 1 till 2 timmar. Frekvensen av mitos i olika vävnader och i olika arter är olika. Till exempel, i den mänskliga röda benmärgen, där 10 miljoner röda blodkroppar bildas varje sekund, bör 10 miljoner mitoser uppstå varje sekund. Och i nervvävnaden är mitoser extremt sällsynta: till exempel i centrala nervsystemet slutar celler i allmänhet att dela sig redan under de första månaderna efter födseln. och i den röda benmärgen, i matsmältningens epitelfoder och i njurrörens epitel, delar de sig till slutet av livet.
Reglering av mitos, frågan om den utlösande mekanismen för mitos.
De faktorer som inducerar cellen till mitos är inte exakt kända. Men man tror att faktorn för förhållandet mellan volymerna i kärnan och cytoplasman (kärnplasma-förhållandet) spelar en viktig roll. Enligt vissa rapporter producerar döende celler ämnen som kan stimulera celldelning. Proteinfaktorerna som var ansvariga för övergången till M-fasen identifierades ursprungligen på basis av cellfusionsexperiment. Fusionen av en cell i vilket stadium som helst av cellcykeln med en cell i M-fasen leder till inträde av kärnan i den första cellen i M-fasen. Detta betyder att det i en cell i M-fasen finns en cytoplasmisk faktor som kan aktivera M-fasen. Senare upptäcktes denna faktor i experiment på överföring av cytoplasma mellan grodaocyter vid olika utvecklingsstadier och utsågs till "mognadsfrämjande faktor" MPF. Ytterligare studie av MPF visade att detta proteinkomplex bestämmer alla M-fashändelser. Figuren visar att sönderdelningen av kärnmembranet, kromosomkondensation, spindelenhet, cytokinese regleras av MPF.
Mitos hämmas av höga temperaturer, höga doser av joniserande strålning och verkan av växtgift. En av dessa gifter kallas colchicine. Med sin hjälp är det möjligt att stoppa mitos i metafasplattans skede, vilket gör det möjligt att beräkna antalet kromosomer och ge var och en av dem en individuell egenskap, dvs att utföra karyotypning.
4. Amitos. Endoreproduktion
Amitos (från grekiska. A - negativ partikel och mitos) -delning av interfaskärnan genom snörning utan att transformera kromosomer. Med amitos finns det ingen enhetlig skillnad mellan kromatider och polerna. Och denna uppdelning föreskriver inte bildandet av genetiskt ekvivalenta kärnor och celler. Jämfört med mitos är amitos en kortare och mer ekonomisk process. Amitotisk uppdelning kan utföras på flera sätt. Den vanligaste typen av amitos är att snöra kärnan i två. Denna process börjar med uppdelningen av kärnan. Förträngningen fördjupas och kärnan är uppdelad i två. Efter detta börjar separationen av cytoplasman, men detta händer inte alltid. Om amitos endast är begränsad till nukleär uppdelning, leder detta till bildandet av bi- och flerkärniga celler. Knoppar och fragmentering av kärnor kan också förekomma med amitos.
En cell som har genomgått amitos kan därefter inte gå in i den normala mitotiska cykeln.
Amitos förekommer i cellerna i olika vävnader från växter och djur. I växter är amitotisk uppdelning ganska vanlig i endospermen, i specialiserade rotceller och i lagringsvävnadsceller. Amitos observeras också i högt specialiserade celler med försvagad livskraft eller degenererande, med olika patologiska processer, såsom malign tillväxt, inflammation, etc.
Huvudprocessen för att förbereda en cell för mitos är DNA-replikering och kromosom duplicering. Men DNA-syntes och mitos är inte direkt relaterade, eftersom den slutliga DNA-syntesen är inte den direkta orsaken till cellens inträde i mitos. I vissa fall, efter att kromosom fördubblats, delar sig cellerna inte, kärnan och alla celler ökar i volym, blir polyploid. Detta fenomen - kromosomreduplicering, utan uppdelning, utvecklades under utvecklingsprocessen som ett sätt att säkerställa tillväxten av organ utan att öka antalet celler. Alla fall där kromosomreduplicering eller DNA-replikering inträffar, men mitos inte förekommer, kallas endoreproduktioner.Cellerna blir polyploid. Som en konstant process observeras endoproduktion i leverceller, epitel i urinvägarna hos däggdjur. När endomitoskromosomer blir synliga efter fördubbling, men kärnkraftshöljet förstörs inte.
Om delande celler kyls ett tag ellerbearbeta dem med alla ämnen som förstör mikrotubulispindel (till exempel kolchicin), så kommer celldelningen att slutaxia. I detta fall kommer spindeln att försvinna och kromosomerna utan avvikelse frånpolerna kommer att fortsätta cykeln av sina omvandlingar: de kommer att börjasvälla, klä med ett kärnkraftsskal. Detta beror påföreningar av alla icke-divergerande uppsättningar kromosomer är storanya kärnor. De kommer naturligtvis initialt att innehålla 4n-numretkromatider och följaktligen 4c mängd DNA. A-priory,det är inte längre en diploid utan en tetraploid cell. Sådan polyplo tycka omceller kan från scenengi gå till S-perioden och, om ta bort kolchicin, dela igen på ett mitotiskt sätt, ger redanavkomma med 4 n antal kromosomer. Som ett resultat kan du fåpolyploidcellinjer med olika ploidivärden. Denna teknik används ofta för att erhålla polyploida växter.
Som det visade sig, i många organ och vävnader med normal diploid organismer från djur och växter möter cellermed stora kärnor, där mängden DNA är multipel2 s.När sådana celler delas upp ser man att antalet kromosomerde har också en multipelökning jämfört med konventionellasom celler. Dessa celler är resultatet av somatiskpolyploidi. Detta fenomen kallas ofta godkänna mat tions- - uppkomsten av celler med ökat DNA-innehåll.Sådana celler uppträder som ett resultat av frånvaroni allmänhet eller ofullständighet i enskilda mitosstadier. Varelsedet finns flera punkter i mitosprocessen, vars blockadkommer att leda till dess stopp och till att polyploidceller uppträder.Blocket kan inträffa under övergången från C2-perioden till rättmen mitos, stopp kan ske i profas och metafas, ii det senare fallet, en kränkning av vägens integritetdivision retena. Slutligen kan abnormiteter i cytotomi också förhindramultiplicera division, vilket kommer att leda till uppkomsten av binukleär och polyploidceller.
Med naturlig blockering av mitos i början, medövergång G 2 - framhäver, celler börjar nästa cykelreplikering, vilket kommer att leda till en progressiv ökning avav DNA i kärnan. I det här fallet, ingen morfologiska särdrag hos sådana kärnor, förutom deras stora storlek.Med en ökning av kärnor detekteras inte mitoti-kromosomer i demav cic-typen. Ofta denna typ av reproduktion utan mitotisk kondenssation av kromosomer förekommer hos ryggradslösa djur, det förekommer också hos ryggradsdjur och växter.Hos ryggradslösa djur, som ett resultat av mitosblockering, är graden av polyploidi kan nå enorma värden. Så i jätteneuroner i blötdjurstritonia, vars kärnor når storlekenupp till 1 mm (!), innehåller mer än 2-10 5 haploida DNA-uppsättningar.Ett annat exempel på en jätte polyploid cell, bildadsom härrör från DNA-reduplicering utan införande av limström till mitos, kan fungera som en silkesekreterande körtelcellsilkesmask. Kärnan har en bisarr förgrening form och kan innehålla stora mängder DNA. Jätteascaris matstrupsceller kan innehålla upp till 100000-talDNA.
Ett speciellt fall av endeproduktion är ökningenploidy av pollenium. Vid polering i S -period under DIC-replikering ny tillsvarta kromosomer fortsätter att förbli despiraliseradestat, men ligger nära varandra, avviker inte ochgenomgår inte mitotisk kondens. I ett sådanti en verkligt interfasform går kromosomerna in i nästa replikationscykel, dupliceras igen och avviker inte. Förbigradvis som ett resultat av replikering och icke-upplösning av kromosomtrådar, en multifilamentös, polytenstruktur av kromosvi är interfaskärnan. Den senare omständigheten är nödvändig undersläpp en linje, eftersom sådana jätte polyten-kromosomernär de inte deltar i mitos, är det dessutom en riktig interfaskromosomer involverade i syntesen av DNA och RNA.De skiljer sig kraftigt från mitotiska kromosomer och i storlekram: flera gånger tjockare än mitotiska kromosomer på grund avsom består av en bunt med flera icke-divergerade kromosmatid - volym polytenkromosomer av Drosophila 1000 gånger “Mer mitotisk. De är 70-250 gånger längre än mitotiskapå grund av det faktum att i kromosomens interfasläge mindre täta (spiraliserade) än mitotiska kromosomer.Dessutom är deras totala antal celler i dipteranerhaploid på grund av att det finns en volym under polytenisering ding, konjugering av homologa kromosomer. Så i Drosophilai en diploid somatisk cell finns 8 kromosomer och i en jättespottkörtelcell - 4.Det finns jätte polyploida kärnor med polyten kromosomer i vissa larver av dipteraner i en burspottkörtlar, tarmar, malpighian-kärl, fettkropp etc. Beskrivna polyten-kromosomer i makronukleus av infusoryster av stilonychia. Denna typ av endoproduktion studeras bäst i insekter.Det har beräknats att i Drosophila cellerna i spottkörtlarnaupp till 6-8 cykler av reduplicering kan förekomma, vilket leder tilltotal cellploidi lika med 1024. I vissa kironomider(deras larver kallas en blodmask) ploidi i dessa celler upp tillnår 8000-32000. I celler börjar polytenkromosomervara synlig efter att ha nått en polyten på 64-128 p, innan detsådana kärnor skiljer sig inte i något annat än storlek från dem runt demdiploida kärnor.
Polytenekromosomer skiljer sig åt i sin struktur: de strukturellt heterogen längd, består av skivor, interdistomter och puffar. Platsritningskivor är strikt karakteristiska för varje kromosom och skiljer sig åtäven i nära besläktade djurarter. Skivor är områden med kondenserad chromatina. Skivorna kan variera i tjocklek. Deras totala antal i polytenkromosomer av kironomider når 1,5-2,5 tusen.Drosophila har cirka 5 tusen skivor.Skivorna är åtskilda av mellanrum, som, precis som skivorna, består av kromatinfibriller, bara lösarepackade. På polytenkromosomer av dipteraner är svullnad ofta synlig,puffar. Det visade sig att puffar förekommer på platser för vissa diskov på grund av deras dekondensering och lossning. I puffarna, avslöjandedet finns RNA, som också syntetiseras där.Mönstret för arrangemang och växling av skivor på polyten-kromosomer är konstant och beror varken på organ eller ålder.djur. Detta är en bra illustration av detsamma kvaliteten på genetisk information i varje cell i kroppen.Puffar är tillfälliga formationer på kromosomer, och under utvecklingen av organismen finns det en viss sekvens i deras utseende och försvinnande på genenolika regioner i kromosomen. Detta efterföljandestyrka är olika för olika tyger. Det har nu bevisats attbildandet av puffar på polyten-kromosomer är ett uttryckgenaktivitet: RNA syntetiseras i puffarna, nödvändigtför att utföra proteinsynteser vid olika stadier av insektsutveckling. Under naturliga förhållanden är Diptera särskilt aktiva ii förhållande till RNA-syntes, de två största puffarna, den så kalladeringarna från Balbiani, som beskrev dem för 100 år sedan.
I andra fall av endoproduktion, WHO-polyploidcellernix som ett resultat av kränkningar av klyvningsapparaten - spindeln:i detta fall inträffar mitotisk kondensation av kromosomer. Sådan fenomenet kallas endomitos,eftersom chro kondensmosome och deras förändringar sker inuti kärnan utan att försvinnakärnkraftsskal.För första gången har fenomenet endomitos studerats väl i celler:olika vävnader i vattenbuggen - - herria. I början av endomikromosomerna kondenseras, på grund av vilka de blir hoväl urskiljbar inuti kärnan, sedan separeras kromatiderna,sträcka ut. Dessa steg, beroende på kromosomernas tillstånd, kan motsvara för att främja profas och metafas av normal mitos. Sedan kromosomerförsvinner i sådana kärnor, och kärnan tar formen av en vanlig interfaskärnan, men dess storlek ökar i enlighet medutvecklingen av ploidi. Efter nästa DNA-reduplikation upprepas denna cykel av endomitos. Som ett resultat,polyploid (32 n) och till och med gigantiska kärnor.En liknande typ av endomitos har beskrivits i utvecklingen av makronukleusugglor i vissa ciliater, i ett antal växter.
Endoreproduktionsresultat: polyploidi och cellförstoring.
Endoreproduktionsvärde: cellens aktivitet avbryts inte. Så till exempel affäreravlägsnande av nervceller skulle leda till deras tillfälliga avstängningfunktioner; endeproduktion tillåter utan avbrott i funktionenöka cellmassan och därigenom öka volymenvi arbetar med en cell.
öka cellproduktiviteten.
Celldelning är central för reproduktion.
Under delningsprocessen uppstår två celler från en cell. Cellen, på basis av assimilering av organiska och oorganiska ämnen, skapar en liknande med en karakteristisk struktur och funktioner.
I celldelning kan två huvudpunkter observeras: uppdelning av kärnan - mitos och uppdelning av cytoplasman - cytokinese, eller cytotomi. Genetikernas huvudsakliga uppmärksamhet är fortfarande nitad på mitos, eftersom ur kromosomteorins synvinkel betraktas kärnan som "ärvets organ".
Under mitos inträffar följande:
- fördubbling av substansen i kromosomer;
- förändringar i det fysiska tillståndet och den kemiska organisationen av kromosomer;
- divergens mellan dotter, eller snarare syster, kromosomer till cellstolparna;
- efterföljande uppdelning av cytoplasman och fullständig restaurering av två nya kärnor i systerceller.
Således innehåller mitos hela livscykeln för kärngener: duplicering, distribution och funktion; som ett resultat av fullbordandet av den mitotiska cykeln, slutar systerceller med lika "arv".
Under delning genomgår cellkärnan fem successiva steg: interfas, profas, metafas, anafas och telofas; vissa cytologer skiljer ett sjätte steg - prometafas.
Mellan två på varandra följande celldelningar är kärnan i mellanfasstadiet. Under denna period har kärnan, när den är fixerad och färgad, en retikulär struktur bildad av färgade tunna trådar, som i nästa fas formas till kromosomer. Även om interfasen kallas annorlunda kärnan i vilofas, på själva kroppen utförs metaboliska processer i kärnan under denna period med störst aktivitet.
Profas är det första steget i att förbereda kärnan för klyvning. I profas förvandlas kärnans retikulära struktur gradvis till kromosomala filament. Från den tidigaste profasen, även under ett ljusmikroskop, kan kromosomernas dubbla natur observeras. Detta antyder att det är i den tidiga eller sena fasen i kärnan som den viktigaste processen med mitos utförs - fördubbling eller reduplikering av kromosomer, där var och en av moderns kromosomer bygger en liknande - dotter. Som ett resultat ser varje kromosom ut i dubbelt längd. Men dessa halvor av kromosomer, som kallas systerkromatider, avvik inte i profas, eftersom de hålls samman av en gemensam plats - centromeren; den centromera platsen delas upp senare. I profas genomgår kromosomer en vridningsprocess längs sin axel, vilket leder till deras förkortning och förtjockning. Det bör betonas att i profas är varje kromosom i karyolymfen lokaliserad slumpmässigt.
I djurceller, även i sen telofas eller mycket tidigt mellanfas, sker en fördubbling av centriolen, varefter konvergensen av dottercentriolerna till polerna och formationerna i astrosfären och spindeln, som kallas en ny apparat, i profasen. Samtidigt upplöses nukleolierna. Ett väsentligt tecken på slutet av profasen är upplösningen av kärnans membran, varigenom kromosomerna är i den totala massan av cytoplasman och karyoplasman, som nu bildar en mixoplasma. Detta avslutar profasen; cellen går in i metafas.
Nyligen, mellan profas och metafas, började forskare skilja mellan ett mellanstadium, kallat prometafas... Prometafas kännetecknas av upplösning och försvinnande av kärnhöljet och rörelse av kromosomer mot cellens ekvatorialplan. Men vid denna tidpunkt har bildandet av achromatinspindeln ännu inte avslutats.
Metafas kallas steget för slutet av kromosomarrangemanget vid spindelns ekvatorn. Det karakteristiska arrangemanget av kromosomer i ekvatorialplanet kallas ekvatorial- eller metafasplattan. Arrangemanget av kromosomer i förhållande till varandra är slumpmässigt. I metafas avslöjas antalet och formen av kromosomer väl, särskilt när man undersöker ekvatorplattan från polerna i celldelningen. Achromatinspindeln är helt formad: spindelfilamenten får en tät konsistens än resten av cytoplasman och fäster vid kromosomens centromerregion. Cytoplasman i cellen under denna period har den lägsta viskositeten.
Anafas Nästa mitosfas kallas, där kromatider delar sig, som nu kan kallas syster- eller dotterkromosomer, och avviker till polerna. I det här fallet avvisas de centromera regionerna från varandra och sedan avviker kromosomerna till polerna. Det måste sägas att divergensen av kromosomer i anafas börjar samtidigt - "som om på kommando" - och slutar mycket snabbt.
I telofasen despiraliseras dotterkromosomerna och förlorar sin synliga individualitet. Kärnans skal och själva kärnan bildas. Kärnan rekonstrueras i omvänd ordning i jämförelse med de förändringar som den genomgick i profas. I slutändan återställs också nukleolierna (eller nukleolerna) och i den mängd i vilken de var närvarande i moderkärnorna. Antalet nukleoler är karakteristiskt för varje celltyp.
Samtidigt börjar cellkroppens symmetriska uppdelning. Kärnorna i dottercellerna passerar in i tillståndet av fas.

Figuren ovan visar ett diagram över cytokinese hos djur- och växtceller. I en djurcell sker uppdelning genom att snöra på modercellens cytoplasma. I en växtcell uppstår bildandet av ett cellseptum när områden med spindelplack bildar ett septum i ekvatorialplanet, kallad phragmoplast. Detta avslutar den mitotiska cykeln. Dess varaktighet beror tydligen på vävnadstypen, organismens fysiologiska tillstånd, externa faktorer (temperatur, ljusförhållanden) och varar från 30 minuter till 3 timmar. Enligt olika författare är passagen för enskilda faser variabel.
Både interna och externa miljöfaktorer som påverkar organismens tillväxt och dess funktionella tillstånd påverkar celldelningens varaktighet och dess individuella faser. Eftersom kärnan spelar en enorm roll i cellens metaboliska processer är det naturligt att tro att längden på de mitotiska faserna kan förändras i enlighet med organvävnadens funktionella tillstånd. Det har till exempel visat sig att under vila och sömn hos djur är den mitotiska aktiviteten hos olika vävnader signifikant högre än under vakenhet. Hos ett antal djur minskar frekvensen av celldelning i ljuset och ökar i mörkret. Det antas också att hormoner påverkar cellens mitotiska aktivitet.
Anledningarna som avgör cellens beredskap att dela sig är fortfarande oklara. Det finns skäl att anta flera sådana skäl:
- fördubblar massan av cellulärt protoplasma, kromosomer och andra organeller, på grund av vilka kärn-plasmaförhållanden störs; för delning måste cellen nå en viss vikt och volym som är karakteristisk för cellerna i denna vävnad;
- fördubbling av kromosomer;
- frisättning av speciella ämnen av kromosomer och andra cellorganeller som stimulerar celldelning.
Mekanismen för kromosomdivergens till polerna i mitafas förblir också oklar. Spindelfilamenten, som är proteinfilament organiserade och orienterade av centrioler och centromerer, spelar uppenbarligen en aktiv roll i denna process.
Arten av mitos, som vi redan har sagt, förändras beroende på vävnadens typ och funktionella tillstånd. Olika typer av mitos är karakteristiska för celler i olika vävnader. I den beskrivna typen av mitos sker celldelning på ett lika och symmetriskt sätt. Som ett resultat av symmetrisk mitos är systerceller ärftliga ekvivalenta när det gäller både kärngener och cytoplasma. Förutom symmetrisk finns det dock andra typer av mitos, nämligen: asymmetrisk mitos, mitos med fördröjd cytokinese, uppdelning av multinukleära celler (uppdelning av syncytia), amitos, endomitos, endoproduktion och polyteni.
När det gäller asymmetrisk mitos är systercellerna ojämna i storlek, mängd cytoplasma och även i förhållande till deras ytterligare öde. Ett exempel på detta är den ojämna storleken på syster- (dotter-) celler från gräshoppan neuroblaster, djurägg under mognad och under spiralklyvning. under uppdelningen av kärnor i pollenkorn kan en av dottercellerna dela sig, den andra inte, och så vidare.
Mitos med fördröjd cytokinese kännetecknas av att cellkärnan delar sig många gånger, och först då delar sig cellkroppen. Som ett resultat av denna uppdelning bildas flerkärniga celler som syncytium. Ett exempel på detta är bildandet av endospermceller och bildandet av sporer.
Amitos direkt klyvning av kärnan utan bildning av klyvningsfigurer kallas. I detta fall inträffar uppdelningen av kärnan genom att "snöra" den i två delar; ibland bildas flera kärnor från en kärna samtidigt (fragmentering). Amitos finns ständigt i cellerna i ett antal specialiserade och patologiska vävnader, till exempel i cancertumörer. Det kan observeras när det utsätts för olika skadliga ämnen (joniserande strålning och höga temperaturer).
Endomitos en sådan process kallas när det sker en fördubbling av kärnklyvning. I detta fall reproduceras kromosomerna, som vanligt, i interfasen, men deras efterföljande divergens inträffar inuti kärnan med bevarande av kärnhöljet och utan bildandet av akromatinspindeln. I vissa fall, även om kärnhöljet löses upp, sker inte separationen av kromosomer till polerna, vilket resulterar i att antalet kromosomer i cellen multipliceras till och med flera tiotals gånger. Endomitos förekommer i cellerna i olika vävnader från både växter och djur. Så, till exempel, visade A.A.Prokofieva-Belgovskaya att genom endomitos i cellerna i specialvävnader: i Cyclops hypodermis, fettkroppen, peritonealt epitel och andra vävnader i fyllningen (Stenobothrus) kan uppsättningen kromosomer öka 10 gånger. Denna multiplikation av antalet kromosomer är associerad med de funktionella egenskaperna hos den differentierade vävnaden.
Under polyeten multipliceras antalet kromosomtrådar: efter reduplikering längs hela längden avviker de inte och förblir intill varandra. I det här fallet multipliceras antalet kromosomfilament inom en kromosom, vilket leder till att kromosomernas diameter ökar markant. Antalet sådana tunna trådar i en polyten-kromosom kan nå 1000-2000. I det här fallet bildas de så kallade jättekromosomerna. Med polyteni faller alla faser av den mitotiska cykeln ut, förutom den viktigaste - reproduktionen av de primära kromosomfilamenten. Fenomenet polyteni observeras i cellerna i ett antal differentierade vävnader, till exempel i vävnaden i spottkörtlarna i dipteraner, i cellerna hos vissa växter och protozoer.
Ibland förekommer en duplicering av en eller flera kromosomer utan några transformationer av kärnan - detta fenomen kallas endoproduktion.
Så, alla faser av mitos i cellen som utgör krävs endast för en typisk process.
i vissa fall, främst i differentierade vävnader, genomgår den mitotiska cykeln förändringar. Cellerna i sådana vävnader har tappat förmågan att reproducera hela organismen, och deras kärnas metaboliska aktivitet är anpassad till den normaliserade vävnadens funktion.
Embryonala och meristemiska celler, som inte har förlorat funktionen att reproducera hela organismen och klassificeras som odifferentierade vävnader, upprätthåller hela mitoscykeln, vilket är grunden för asexuell och vegetativ reproduktion.
Om du hittar ett fel, välj en textbit och tryck på Ctrl + Enter.
Sekvensen av faser i den mitotiska cykeln visas i fig. 4.
Figur: 4. Faser av mitos
Prophase. I profas ökar kärnan, och kromosomala filament, som vid denna tidpunkt redan är spiraliserade, blir tydliga.
Varje kromosom består, efter reduplikering i interfas, av två systerkromatider förbundna med en centromer. I slutet av profasen försvinner vanligtvis kärnhöljet och nukleolierna. Ibland försvinner kärnan i nästa mitosfas. Man kan alltid hitta tidiga och sena betoning på förberedelserna och jämföra dem med varandra. Förändringarna är tydligt synliga: kärnan och kärnans skal försvinner. Kromosomala trådar är tydligare i det sena profaset, och det kan ofta ses att de dupliceras. I profas finns det också en divergens mellan centriolerna, som bildar två poler i cellen.
Prometafas börjar med den snabba upplösningen av kärnhöljet i små fragment som inte kan särskiljas från fragment av det endoplasmiska nätverket (fig. 5). I kromosomer, på varje sida av centromeren i prometafas, bildas speciella strukturer, kallade kinetokorer. De fäster vid en speciell grupp mikrotubuli som kallas kinetochore-filament eller kinetochore mikrotubuli. Dessa trådar sträcker sig från båda sidor av varje kromosom, går i motsatta riktningar och interagerar med filamenten i den bipolära spindeln. I det här fallet börjar kromosomerna röra sig intensivt.

Figur: 5. Prometafas (figuren av moderstjärnan är byggd) i en pigmenterad cell. Heidenhain-järnhematoxylinfärgning. Genomsnittlig ökning
Metafas. Efter att kärnhöljet försvinner kan man se att kromosomerna har nått maximal spiralisering, blir kortare och rör sig till cellens ekvatorn och ligger i samma plan. Centriolerna som ligger vid cellens poler kompletterar bildningen av delningsspindeln och dess filament fäster vid kromosomerna i centromerregionen. Centromererna i alla kromosomer är i samma ekvatorialplan och axlarna kan placeras ovanför eller under. Denna kromosomposition är bekväm för att räkna och studera morfologi.
Anafas börjar med sammandragningen av trådarna i spindeldelningen, på grund av vilken uppstår kan placeras ovanför eller under. Allt detta är bekvämt för att räkna antalet kromosomer, studera deras morfologi och uppdelning av centromerer. I mitosens anafas delas den centromera regionen i var och en av de två-kromatidkromosomerna, vilket leder till separationen av systerkromatider och deras transformation till oberoende kromosomer (det formella förhållandet mellan antalet kromosomer och DNA-molekyler är 4n4c).
Detta är den exakta fördelningen av det genetiska materialet, och vid varje pol finns det samma antal kromosomer som den ursprungliga cellen hade innan de fördubblades.
Förflyttningen av kromatider till polerna sker på grund av sammandragningen av sträckfilamenten och förlängningen av de stödfibrerna i den mitotiska spindeln.
Telofas. Efter avslutad divergens av kromosomer till modercellens poler bildas två dotterceller i telofasen, som var och en får en fullständig uppsättning av enstaka kromatidkromosomer i modercellen (formel 2n2c för var och en av dottercellerna).
I telofas genomgår kromosomerna vid varje pol despiralisering, dvs. en process motsatt den som sker i profas. Kromosomernas konturer förlorar sin klarhet, den mitotiska spindeln förstörs, kärnhöljet återställs och nukleolerna dyker upp. Uppdelningen av cellkärnor kallas karyokinesis (fig. 6).
Därefter bildas en cellvägg från phragmoplasten, som delar hela innehållet i cytoplasman i två lika stora delar. Denna process kallas cytokinese. Det här är hur mitos slutar.

Figur: 6. Faser av mitos i olika växter

Figur: 7. Fördelning av homologa kromosomer och gener som finns i dem under den mitotiska cykeln i en hypotetisk organism (2n \u003d 2) generationer och genetisk livskontinuitet vid asexuell reproduktion av organismer.
Grundläggande termer och begrepp: anafas; dottercell; mellanfas; mor (förälder) cell; metafas; mitos (period M); mitotisk (cell) cykel; postsyntetisk period (G 2); presyntesperiod (G1); profas; systerkromatider; syntetisk period (S); telofas; kromatid; kromatin; kromosom; centromer.
-
 Intima-media-komplex: diagnostiskt värde Mer tät i intensitet
Intima-media-komplex: diagnostiskt värde Mer tät i intensitet
-
 Hur man behandlar blodkärl vid diabetes mellitus - kontraindikationer och rekommendationer Behandling av blodkärl i benen vid diabetes mellitus
Hur man behandlar blodkärl vid diabetes mellitus - kontraindikationer och rekommendationer Behandling av blodkärl i benen vid diabetes mellitus
-
 Tea tree eterisk olja: egenskaper, applikationer, recensioner Tea tree oil från sprickor applikation
Tea tree eterisk olja: egenskaper, applikationer, recensioner Tea tree oil från sprickor applikation
-
 Blodtransfusionskomplikationer
Blodtransfusionskomplikationer